
Ceci est une version archivée de Batary2008Francais à 2010-09-02 09:47:58.
Distribution des ufs de la Diane Zerynthia polyxena (Lepidoptera, Papilionidae)
par P. BATÁRY (1,2), N. ÖRVÖSSY (1), Á. KŐRÖSI (3) and L. PEREGOVITS (1), 2008. In Acta Zoologica Academiae Scientiarum Hungaricae, 54 (4) : 401-410.
- 1 Department of Zoology, Hungarian Natural History Museum H-1088 Budapest, Baross u. 13, Hungary
- 2 Current address: Agroecology, Georg-August University Waldweg 26, D-37073 Göttingen, Germany
- 3 Animal Ecology Research Group of the Hungarian Academy of Sciences and the Hungarian Natural History Museum, H-1083 Budapest, Ludovika tér 2, Hungary
RÉSUMÉ
Nous avons étudié les facteurs environnementaux qui influencent, à différentes échelles, la répartition spatiale des ufs d'un papillon, la Diane Zerynthia polyxena sur son unique plante-hôte l'Aristoloche clématite Aristolochia clematitis [en Hongrie]. Deux ensembles de variables ont été considérées : la nature de l'habitat (plantations de Robinier faux-acacia, clairières, merlons) et la répartition des plantes-hôtes. Nous avons enregistré la distribution des ufs sur les plantes. Nous avons compté enregistré l'évolution de la végétation autour des pontes. Nous avons également mesuré la taille des feuilles et compté le numéro des feuilles sur chaque plant portant des pontes. L'apparence des pieds à été caractérisée en fonction de la différence de hauteur entre les tiges portant des ufs et les tiges périphériques. Deux variables ont été enregistrées sur les pontes (importance de la nourriture / taille des pieds) : nombre de feuilles et emplacement des ufs dans l'ordre des feuilles.
Le type d'habitat affecte la distribution des ufs : les plantations de Robinier faux-acacia et les merlons sont plus attractifs que les clairières. À une échelle plus fine, la densité ou laspect général de la plante-hôte nont pas dincidence sur la distribution des ufs. Au niveau des plantes, le nombre d'ufs augmente avec le nombre de feuilles sur chaque pied, et la position des ufs est relatif au nombre d'ufs par pieds : un nombre d'ufs réduit implique des pontes plus hautes sur la plante. Les acteurs concluent que les relations intervenant dans la distribution spatiales des ufs ne peuvent être correctement interprétées quen lien avec une analyse hiérarchisée des facteurs environnementaux.
Mots-clés
Zerynthia polyxena, oviposition, intéraction planteinsecte, Aristolochia clematitis, plante-hôte, échelle spatiale
INTRODUCTION
La ponte est une intéraction écologique particulièrement importante entre les insectes phytophages et leurs plantes-hôtes (RABASA et al. 2005). De plus, les préférences dans le choix du support de ponte et donc du développement larvaire sont au centre des relations biologiques entre l'insecte et la plante (XUE et al. 2007). Les imagos femelles sont capables de faire la différence entre différents sites de pontes, en fonction de leurs régimes climatiques, de leurs qualité d'alimentation pour les chenilles, ou des niveaux potentiels de concurrence et de prédation (BERNARDO 1996). La variation de ces aspects environnementaux affecte ainsi la performance de production d'une descendance. Chez les papillons, la ponte peut présenter des préférences envers certaines espèces, en particulier pour certaines en particulier, voire certaines parties des plante-hôtes, ce qui permet de déterminer les caractéristiques physiques et chimiques auxquels les insectes réagissent (THOMPSON & PELLMYR 1991, BERNAYS&CHAPMAN 1994). Etant donné que les chenilles émergentes sont relativement immobiles, la clé de leur survie et leur développement réside dans le choix de l'emplacement de la ponte par la femelle (PORTER 1992). En dehors d'études expérimentales (e.g. SINGER et al. 1993), il existe deux méthodes répandues d'études des sites de pontes des femelles de papillons. L'une d'entre-elles est basée sur le suivi des femelles et de l'observation visuelle de l'oviposition (e.g. GRUNDEL et al. 1998, BERGMAN 1999, ZIMMERMANN et al. 2005, KŐRÖSI et al. 2008). L'autre méthode consiste à observer la présence-absence ou encore la distribution visibles des ufs sur les plantes-hôtes (e.g. FLOATER&ZALUCKI 2000, ELLIS 2003, RABASA et al. 2005). Cette méthode indirecte est plus souple car elle permet au chercheur de planifier son plan d'échantillonnage et d'obtenir un échantillon beaucoup plus grand (sous réserve d'une certaine visibilité et de la possibilité d'identification des ufs). Cependant, l'inconvénient évident de cette méthode par rapport à la première, c'est que l'on ne tient compte que des plantes-hôtes habituellement utilisées par le papillon (DE BOER & HANSON 1984).
Plusieurs études ont testé les facteurs influençant la distribution des ufs à différences échelles spatiales. MCKAY (1991) a étudié les exigences de ponte du Citron Gonepteryx rhamni dans des boisements humides, en tenant compte de la distribution spatiale des plantes-hôtes, des facteurs physico-chimiques et a trouvé que la plupart des ufs étaient pondus sur de jeunes arbres poussant dans les endroits ensoleillés. De plus, les papillons semblaient préférer les arbres avec un sous-bois peu dense. De même, FLOATER and ZALUCKI (2000) ont étudié la qualité des arbres-hôtes et des préférences de la mite Australienne Ochrogaster lunifer. Ils ont trouvé plus d'amas d'ufs sur les arbres de grande qualité dans les habitats ouverts mais homogènes, tandis que dans les habitats présentant des boisement mixtes, les pontes occupaient des arbres de moins grande qualité. Dans le cas de la ponte du Collier des hélianthèmes Aricia artaxerxes ELLIS (2003), a décrit les effets
Il a décrit les incidences des différentes plantes-hôtes sur les pontes en fonctions des facteurs du micro-environnement, comme le niveau de couverture du sol par la plante-hôte ou la hauteur de végétation. Les ufs de ce papillon ont été plus fréquents sur les feuilles des plantes-hôtes les plus jeunes et plus et plus dans les sites non gérés que dans les sites gérés (végétation plus courte), tandis que la densité de plantes-hôtes ou le niveau de recouvrement du sol n'a pas d'incidence sur les pontes. Par ailleurs, FARTMANN (2006) a étudié les effets des plantes-hôtes, du micro-environnement et du micro-climat sur la distribution des ufs de la Lucine Hamearis lucina. Il a découvert des ufs à une hauteur moindre sur les plantes-hôtes (Primula veris) et une majorité de pontes sur les stations recevant une insolation directe entre 09:00 à 17:00 et où le couvert herbacé dépassait 60% de recouvrement. DENNIS (1996) a étudié la ponte de Zerynthia cretica en fonction des feuilles de plantes-hôtes, des stades de croissance et des touffes. Il trouva que les femelles pondaient plus d'ufs sur les touffes les plus grosses et présentant des feuilles plus larges, en particulier sur les brins extérieurs des touffes. Nous avons seulement trouvé deux études qui se sont focalisées sur des papillons monophages pour lesquels ils ont utilisé la méthode de présence-absence des ufs pour tester des facteurs à différentes échelles spatiales, selon des modèles hiérarchiques emboîtés. KÉRY et al. (2001) a étudié la présence des ufs de Maculinea rebeli sur les fruits de pieds et de populations de Gentiana cruciata et a observé que les facteurs mesuré au niveau des pieds étaient plus importants que ceux mesurés à l'échelle de la population. RABASA et al. (2005) a utilisé modèle imbriqué similaire pour détecter la présence d'ufs de Iolana iolas sur les pieds et sur les stations de l'arbuste Colutea hispanica/ / en fruits. Ils montrent que des facteurs importants infuencent la présence des ufs à chaque échelle. Dans la présente étude, nous avons essayé de comprendre quels sont les facteurs environnementaux qui influencent le plus la densité et la distribution des ufs de Z. polyxena dans les mosaïques de plantations de Robinier faux-acacia et de peupliers, dans lesquelles la plante-hôte du papillon est abondante.
MATERIEL ET MÉTHODE
Espèce étudiée
La Diane Zerynthia polyxena DENIS et SCHIFFERMÜLLER, 1775 est un papilionidé qui atteind le nord de son aire de répartition en Europe Centrale (TOLMAN 1997). L'espèce est protégée réglementairement en Hongrie. C'est un papillon monophage en Hongrie, se nourrissant sur une plante herbacée, l'Aristoloche clématite Aristolochia clematitis LINNEAUS, 1753, (Aristolochiaceae). Cette plante-hôte est commune dans les habitats anthropisés, comme les plaines inondables, les vergers, les bords de routes, ou les plantations de Robinier faux-acacia Robinia pseudoacacia LINNEAUS, 1753 (Fabaceae) et de peuplier hybride Populus × euramericana (Salicaceae). La période de vol du papillon commence à la mi/fin avril et se termine à la mi/fin mai. Les femelles pondent sur la face supérieure des feuilles, un uf isolé, un petit amas ou un grand amas d'ufs. Contrairement à Zerynthia rumina LINNEAUS, 1758, qui possède deux plantes-hôtes (Aristolochia baetica LINNEAUS, 1753, Aristolochiaceae and Aristolochia longa LINNEAUS, 1753, Aristolochiaceae), Z. polyxena est seulement monophage en Hongrie ; les ufs et les chenilles sont seulement observés sur A. clematitis (ROTHSCHILD et al. 1972, JORDANO&GOMARIZ 1994,ÖRVÖSSY et al. non publ.)
Aire d'étude et méthode d'échantillonnage
L'étude a été conduite dans la grande plaine hongroise près de Csévharaszt (Central Hungary, 47°18N, 19°26E), dans un territoire de plantations, principalement de Robinier et de Peupliers interrompues par des clairières. Les 0,02 à 0,03 km2 de parcelles sont séparées par des buttes résultant de la dernière exploitation. Ces buttes sont composées de souches et de racines recouvertes de terre et constituant des stations idéales pour l'Aristoloche clématite. Une touffe de plantes-hôtes comprenant au moins cinq tiges par m2 et espacées d'au moins 10m de la touffe suivante a été considérée comme une station. Nous avons choisi 4x4 stations pour notre échantillonnage dans les trois types d'habitat : dans les plantations de Robinier, dans les clairières et sur les merlons. Dans notre précédente étude dans le même territoire, nous avions observé que les imagos évitaient les plantations de peupliers (ÖRVÖSSY et al. 2005). Chacune des stations de plantes-hôtes ont été occupés par plusieurs milliers de pousses aristoloche. Nous choisis au hasard 10 points au sein de chaque station, où nous avons vérifié la présence d'ufs sur les pousses de la plante-hôte dans un cercle de 5 mètres de rayon. Dès que quelques pousses d'Aristoloches présentaient des feuilles portant des ufs, nous arrêtions les recherches, après avoir constaté le premier pied avec des ufs. Ces cercles ont été couverts par une moyenne de 775 pousses d'Aristoloches. Sur un total de 120 cercles choisis au hasard, 98 contenaient des ufs, et les cercles sans ufs ont été exclus des analyses. La période de vol (du 26 avril au 15 mai) s'est achevée avant la recherche des ufs (du 17 au 23 mai). Ainsi de nouvelles pendant la période d'échantillonnage n'étaient plus possible, ce qui empêcherait tout biais éventuel. Aucune chenille n'a été trouvée lors de la cadre de la recherche des ufs.
Au cours de la période d'échantillonnages, plusieurs variables environnementales ont été mesurées à proximité immédiate du site d'étude. Nous avons regroupé ces variables en fonction d'échelles spatiales. Le type d'habitat d'une station (plantation de robiniers, clairières ou merlon) a été considéré comme le premier niveau de variable et incorporé dans le modèle comme l'un des facteurs. Pour caractériser les types d'habitats, nous avons compté la densité de plantes-hôtes dans 20 relevés de 4m2 dans chaque station. Nous avons mesuré la taille des stations. Pour caractériser le micro-environnement de chaque relevé d'Aristoloches, nous avons comptabilisé le nombre de stations et mesuré la taille moyenne des plantes dans un carré de 1m de côté. Nous avons également mesuré la taille et compté les feuilles des pieds portant des ufs dans chaque station. L'apparence des pieds à été caractérisée en fonction de la différence de hauteur entre les tiges portant des ufs et les tiges périphériques dans un carré de 1m de côté. Sur les pieds portant les ufs, deux variables ont été mesurées : le nombre de feuilles par pied et la position des ufs ou de la grappe d'oeufs (l'échelle de ces variables donne le niveau d'aliments de la plante, à ne pas confondre avec l'échelle des variables mesurées dans la proximité directe des pousses portant des ufs qui informe sur le micro-environnement). La dernière variable correspond au nombre de feuilles sous la feuille portant les ufs, divisés par le nombre total de feuilles.
L'analyse statistique
Les effets des variables mentionnées ci-dessus sur la distribution des ufs ont été analysés à travers des modèles généraux linéaires à effets mixtes avec la méthode dite "Restricted Maximum Likelihood" (vraisemblance restreinte maximale). La normalité de la distribution des ufs sur des feuilles a été évaluée en utilisant des parcelles normales divisées en quantiles. Une conversion logarithmique a été appliquée à la distribution non-normale. Les variables suivantes, non corrélées ont été considérées dans le modèle statistique :
- 1) le type d'habitat en tant que facteur et la taille de stations en tant que co-variable au niveau de stations,
- 2) au niveau du micro-environnement, la densité des tiges d'Aristoloches autour de la pousse portant des ufs et leur apparence ;
3) le nombre de feuilles d'aristoloches et la position des ufs au niveau de la plante. Comme les plantes-hôtes sont incluses dans les stations, ce dernier a été utilisé comme un facteur aléatoire. Bien que les feuilles d'Aristoloches soient également incluses dans les plantes-hôtes, nous ne pouvions pas inclure les plantes-hôtes comme un facteur aléatoire dans le modèle, car cela aurait provoqué un effet trop important.
Au total, on obtenait des ufs sur deux feuilles pour seulement cinq pousses de plantes-hôtes et des ufs sur trois feuilles pour une seule plante-hôte. Les feuilles sans ufs (1036) ont été exclues de l'analyse. De plus, nous avons comparé les modèles avec et sans l'inclusion de la plante-hôte comme un facteur aléatoire. Nous avons constaté que les deux modèles ont été significativement différents (L-ratio = 59,8, p <0,0001), et le modèle sans plante-hôte obtenait une plus petite valeur d'AIC, ce qui indique que ce modèle était plus robuste. Par conséquent, nous avons décidé de n'utiliser que ce dernier modèle. Les calculs ont été effectués en utilisant R (R Development Core Team 2006, version 2.2.1) et la configuration nlme R (PINHEIRO et al. 2007, version 3.1).
RESULTS
Nous avons comptabilisé 597 ufs de Z. polyxena pondus isolément, ou en petit (2 à 8 ufs) ou en gros amas (10 à 99 ufs) posés à la surface supérieure des feuilles. Au cours des investigations, nous n'avons pas trouvé de chenilles ni d'ufs éclos. Le type d'habitat affecte significativement la distribution des ufs (tableau 1). Lorsque nous avons comparé la densité des pieds d'Aristoloches entre les types d'habitat avec une ANOVA à sens unique, nous avons trouvé des différences significatives (F = 23,207, p <0,001, N = 240, Fig. 1. Les tests Tukey HSD post-hoc ont montré que tous les trois types d'habitats diffèrent sensiblement les uns des autres en ce qui concerne la densité de pousses d'Aristoloches.
Il y avait beaucoup plus d'ufs sur les Aristoloches dans les plantations de Robinier et sur les merlons que dans les clairières (Fig. 2). Au niveau micro-environnement, ni la densité des pousses d'Aristoloches, ni l'apparence des Aristoloches n'affecte la distribution des ufs (tableau 1). A l'échelle des pieds d'Aristoloches, le nombre d'ufs augmente de manière significative avec le nombre de feuilles par pied (tableau 1). Enfin, la position des ufs dépend également de façon significative du nombre d'ufs ; il y avait plus d'ufs sur la partie inférieure de l'Aristoloche que vers le haut (Fig. 3).
Tableau 1. Modèles linéaires mixtes permettant de tester la détermination des préférences de ponte de Z. Polyxena à différents niveaux. Les valeurs p en gras indiquent des effets importants.
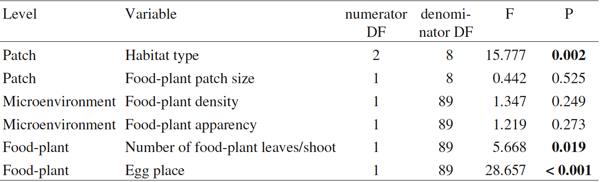
Fig. 1. Moyenne (± SE) du nombre de pousses d'Aristoloches par m2 dans les trois types d'habitats. Les différentes lettres indiquent des différences significatives à P <0,05
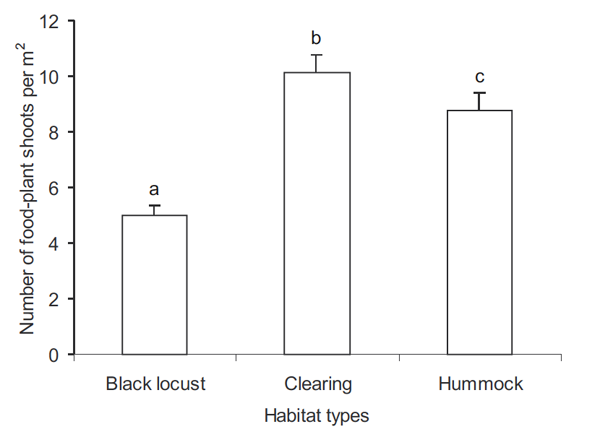
Fig. 2. Moyenne (±SE) du nombre d'ufs de Z. polyxena par feuille d'Aristoloche dans les trois types d'habitats. Les différentes lettres indiquent des différences significatives à P <0,05
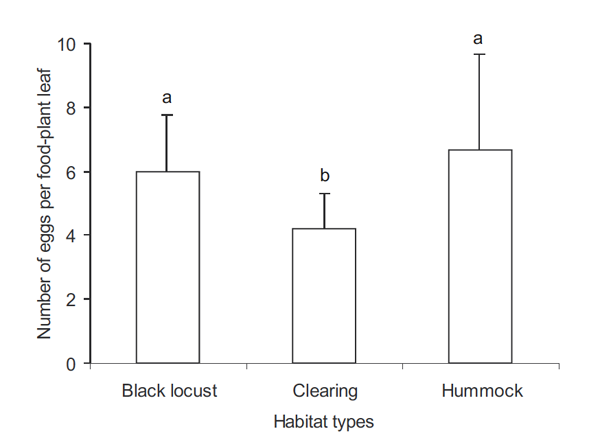
Fig. 3. Position des ufs sur les feuilles par pied (le nombre de feuilles moins les feuilles portant les ufs divisé par le nombre total de feuilles) est portée en log du nombre d'ufs. Une valeur faible de la position des ufs indique que les ufs sont sur les feuilles inférieures. La ligne continue représente la position moyenne des ufs dans le modèle ajusté.
DISCUSSION
Le comportement conduisant à la ponte, à savoir la sélection du site de ponte, est un processus complexe, car une ponte réussie contribue grandement à la production d'une descendance (SCHOWALTER 2006). Cependant, les femelles ne sélectionnent pas toujours la plante-hôte la plus appropriée et les chenilles nouvellement écloses peuvent rejeter la plante sur laquelle elles éclosent (BERNAYS & Chapman, 1994). Les pontes aberrantes peuvent être partiellement corrigées par les chenilles, mais si elles veulent survivre, la préférence des femelles et la performance des chenilles doivent se chevaucher autant que possible. Dans la présente étude, la distribution des ufs de Z. polyxena a été affectée sur deux niveaux, essentiellement par le type d'habitat et aussi par les caractéristiques au niveau des plantes-hôtes, mais pas par des facteurs au niveau micro-environnemental.
Les différences d'habitat rencontrées dans la présente étude, à savoir la plantation de Robinier et des merlons sont plus privilégiées que les clairières, pourrait bien être interprétée comme la préférence des papillons évitant les sites les plus exposés aux rayons directs du soleil ou au froid. FARTMANN (2006) a signalé que la Lucine préférait les sites de ponte localisées en prairies calcaires, où les plantes-hôtes (Primula veris) reçoivent un ensoleillement direct entre 09h00 et 17h00. MCKAY (1991) a constaté que la plupart des ufs de G. rhamni ont été pondus sur de petits arbres isolés (Frangula alnus) poussant dans les secteurs les plus ensoleillés. Ces études montrent que les préférences de sites de ponte correspondent à ceux dont le microclimat serait optimal pour le développement des chenilles.
Dans notre cas, la taille des parcelles n'a pas d'incidence sur le nombre d'ufs pondus, tandis que dans le cas de Z. cretica DENNIS (1996) a montré que les taches de plantes-hôtes les plus grandes présentaient plus d'ufs. Toutefois, nous devons appeler l'attention sur le fait que, dans cette étude, les stations avaient été cartographiés à une échelle beaucoup plus fine (taille des stations d'Aristoloches : de 25 à 2400 cm2) que dans notre cas (intervalle de la dimension des stations : de 661 à 11954 m2, avec une moyenne de 3384 m2). Au niveau micro-environnemental, ni la densité des pousses d'Aristoloches, ni l'aspect des plantes-hôtes ne semble agir comme un facteur limitant la densité des ufs. Toutefois, nous devons mentionner que les pousses d'Aristoloches sont présentes en très grand nombre dans chaque habitat. En outre, la densité des pousses d'Aristoloches était plus élevé dans les clairières, généralement deux fois plus élevé que dans les plantations de Robinier et environ 15% de plus que sur les merlons (Fig. 1). Cela suggère également que l'incidence de l'habitat ne dépend pas de la disponibilité des plantes-hôtes. Contrairement à nos résultats, DENNIS (1996) a trouvé dans son étude fine que le nombre de pieds d'Aristoloches a été l'un des facteurs les plus importants affectant la densité des ufs de Z. cretica. En ce qui concerne l'aspect des végétaux, la hauteur des pousses portant les ufs était significativement plus élevée que ceux dans le micro-environnement (t-test apparié, t = 8,956, P <0,001), cela influence probablement le choix des plantes-hôtes, mais certainement pas la charge d'ufs.
Le nombre de feuilles d'Aristoloches a montré un effet positif sur le nombre d'ufs et plus d'ufs ont été trouvés alors sur les feuilles du bas des tiges. Dans notre étude antérieure, nous avons observé que, au début de la période de vol les plantes-hôtes étaient très rares et commençaient à peine à se développer. Par conséquent, les premières femmes avaient un choix restreint pour pondre (ÖRVÖSSY et al. 2005). Cela signifie probablement qu'une repousse précoce d'Aristoloches présente un certain avantage par rapport à une repousse tardive, car elles seraient plus apparentes pour les papillons. Lorsque les pousses reçoivent des ufs à un stade plus jeune, les ufs se trouvent sur les feuilles inférieures. En outre, à cette époque, les femmes ont encore une charge importante d'ufs, ce qui pourrait provoquer des pontes numériquement plus importantes (en grappes). Cet aspect suggère que les données temporelles sont également importants à prendre en compte pour mieux comprendre la ponte. DENNIS (1996) a également souligné l'importance de la durée de développement des Aristoloches, il soupçonnait que les stations d'Aristoloches se développant sur une période plus longue étaient susceptibles d'avoir plus de chenilles.
Travaillant sur une espèce proche, Z. rumina, JORDANO & GOMARIZ (1994) observèrent que les chenilles fraîchement écloses consommaient les feuilles plus jeunes et les plus tendres de la plante-hôte. Cela pourrait indiquer que les femelles de Z. polyxena pondent en priorité sur de jeunes feuilles. Cependant, à partir d'une recherche sur les ufs, il n'était pas possible d'enquêter sur d'éventuels effets temporels. Une autre explication possible pour la sélection de jeunes feuilles serait que les femelles sélectionnent ces feuilles en raison d'une faible concentration de défences chimiques. Toutefois, ROTHSCHILD et al. (1972) décrit que Z. polyxena contient et assimile de manière efficace deux acides aristolochiques qui sont probablement présents dans sa plante-hôte, A. clematitis .
A travers cette courte étude, nous avons montré que la distribution des ufs de Z. polyxena est déterminé par plusieurs éléments agissant à différents niveaux. Nous devons souligner que le processus complexe, tel que la ponte, pourrait également être étudié par le suivi direct des femmes, ce qui permettrait d'éclairer d'autres aspects de la distribution des eufs. En conclusion, nous soulignerons que les études concernant la distribution spatiale et corrélée des ufs de papillons doivent être analysés en considérant une structure hiérarchisée et intrinsèque des facteurs environnementaux (RABASA et al. 2005).
Acknowledgements
Deux personnes anonymes nous ont transmis des remarques constructives qui ont considérablement augmenté le manuscrit. Nous sommes reconnaissant à LAJOS RÓZSA pour ses nombreux commentaires et la révision linguistique du manuscrit. L'étude a été conduite dans le cadre du programme national recherche & développement intitulé The origin and genezis of the fauna of the Carpathian Basin: diversity, biogeographical hotspots and nature conservation significance (contract no. 3B02304).
REFERENCES
BERGMAN, K.-O. (1999) Habitat utilization by Lopinga achine (Nymphalidae: Satyrinae) larvae and ovipositing females: implications for conservation. Biological Conservation 88: 6974.
BERNARDO, J. (1996) Maternal effects in animal ecology. American Zoologist 36: 83105.
BERNAYS, E. A. & CHAPMAN, R. F. (1994) Host-plant selection by phytophagous insects. Chapman & Hall, London, 312 pp.
BOER, DE G. & HANSON, F. E. (1984) Foodplant selection and induction of feeding preference among host and non-host plants in larvae of the tobacco hornworm Manduca sexta. Entomologia Experimentalis et Applicata 35: 177193.
DENNIS, R. L. H. (1996) Oviposition in Zerynthia cretica (Rebel, 1904): loading on leaves, shoots and plant patches (Lepidoptera, Papilionidae). Nota Lepidopterologica 18: 315.
ELLIS, S. (2003) Habitat quality and management for the northern brown argus butterfly Aricia artaxerxes (Lepidoptera: Lycaenidae) in North East England. Biological Conservation 113: 285294.
FARTMANN, T. (2006) Oviposition preferences, adjacencies of old woodland and isolation explain the distribution of the Duke of Burgundy butterfly (Hamearis lucina) in calcareous grasslands in central Germany. Annales Zoologici Fennici 43: 335347.
FLOATER, G. J.&ZALUCKI,M. P. (2000) Habitat structure and egg distributions in the processionary caterpillar Ochrogaster lunifer: lessons for conservation and pest management. Journal of Applied Ecology 37: 8799.
GRUNDEL, R., PAVLOVIC, N. B. & SULZMAN, C. L. (1998) Habitat use by the endangered Karner blue butterfly in oak woodlands: the influence of canopy cover. Biological Conservation 85: 4753.
JORDANO, D. & GOMARIZ, G. (1994) Variation in phenology and nutritional quality between host plants and its effect on larval performance in a specialist butterfly, Zerynthia rumina. Entomologia Experimentalis et Applicata 71: 271277.
KÉRY, M., MATTHIES, D. & FISCHER, M. (2001) The effect of plant population size on the interactions between the rare plant Gentiana cruciata and its specialized herbivore Maculinea rebeli. Journal of Ecology 89: 418427.
KŐRÖSI, Á., ÖRVÖSSY, N., BATÁRY, P., KÖVÉR, S. & PEREGOVITS, L. (2008) Restricted within habitat movement and time-constrained egg-laying in female Maculinea rebeli butterflies. Oecologia 156(2): 455464.
MCKAY, H. V. (1991) Egg-laying requirements of woodland butterflies; Brimstones (Gonepteryx rhamni) and Alder Buckthorn (Frangula alnus). Journal of Applied Ecology 28: 731743.
ÖRVÖSSY, N., KŐRÖSI, Á., VOZÁR, Á., BATÁRY, P. & PEREGOVITS, L. (2005) Microhabitat preference of the Southern Festoon (Zerynthia polyxena). P. 24. In: SETTELE, J., KÜHN, E. & THOMAS, J. A. (eds): Studies on the ecology and conservation of butterflies in Europe. Vol. 1. General concepts and case studies. PENSOFT Publishers, Sofia.
Pinheiro200è PINHEIRO, J., BATES, D., DEBROY, S. & DEEPAYAN, S. (2007) The nlme Package: Linear and Nonlinear Mixed Effects Models. URL: http://cran.r-project.org/src/contrib/Descriptions/nlme.html
PORTER, K. (1992) Eggs and egg-laying. Pp. 4672. In: DENNIS, R. L. H. (ed.) The ecology of butterflies in Britain. Oxford University Press, Oxford.
RABASA, S. G., GUTIÉRREZ, D. & ESCUDERO, A. (2005) Egg laying by a butterfly on a fragmented host plant: a multi-level approach. Ecography 28: 629639.
R DEVELOPMENT CORE TEAM (2006) R: a language and environment for statistical computing.
Foundation for Statistical Computing, Vienna. URL: http://www.R-project.org
ROTHSCHILD,M., EUW, J. VON&REICHSTEIN, T. (1972) Aristolochic acids stored by Zerynthia polyxena (Lepidoptera). Insect Biochemistry 2: 334343.
SCHOWALTER, T.D. (2006) Insect ecology. An ecosystem approach. Academic Press, London, 572 pp.
SINGER, M. C., THOMAS, C. D. & PARMESAN, C. (1993) Rapid human-induced evolution of insecthost associations. Nature 366: 681683.
THOMPSON, J. N. & PELLMYR, O. (1991) Evolution of oviposition behavior and host preference in Lepidoptera. Annual Review of Entomology 36: 6589.
TOLMAN, T. (1997) Butterflies of Britain and Europe. Collins Field Guide. HarperCollins , London, 320 pp.
XUE, H.-J., EGAS, M. & YANG, X.-K. (2007) Development of a positive preferenceperformance relationship in an oligophagous beetle: adaptive learning? Entomologia Experimentalis et Applicata 125: 119124.
ZIMMERMANN, K., FRIC, Z., FILIPOVÁ, L. & KONVIČKA, M. (2005) Adult demography, dispersal and behaviour of Brenthis ino (Lepidoptera: Nymphalidae): how to be a successful wetland butterfly. European Journal of Entomology 102: 699706.
Traduction proposée par Christophe Bernier, le 1er septembre 2010.
Il n'y a pas de commentaire sur cette page.
[Afficher commentaires/formulaire]


{kind=link}
{kind=link}
{kind=link}
{kind=link}